Write as second assignment
Stelar Evolution in Pteridophytes
Stelar Evolution in Pteridophytes
Central cylinder or core of vascular tissue consisting of xylem, phloem, pericycle and sometimes medullary rays and pith is called Stele.
The word stele is derived from Greek which means central pillar. The Stele is the central cylinder and is separated from the cortex by the endodermis. The endodermis is the innermost layer of the cortex and pericycle is the outermost portion of the Stele.
Van Tiegam and Douliot proposed Stellar Theory in 1886.
According to to the the stellar theory there is no fundamental difference in the gross anatomy of the stem and root because in both of them is Stele surrounded by cortex is present
According to this theory the stele is not only made up of xylem and phloem but the tissue like pericycle, vascular rays and pith are also associated with it.
According to to the the stellar theory there is no fundamental difference in the gross anatomy of the stem and root because in both of them is Stele surrounded by cortex is present
According to this theory the stele is not only made up of xylem and phloem but the tissue like pericycle, vascular rays and pith are also associated with it.
Types of steles
Easu and Smith recognise two principal types of stelar organisation among the vascular plants. These are the Protostele and Siphonostele.
Protostele
The name protostele was suggested by Jeffrey and he regarded it as the most primitive and simplest type of stele.
Steel in which the gas cylinder consists of a solid core of xylem surrounded by phloem pericycle and endodermis is called protostele. There is no pith in a protostele. it is a fundamental type of Steel of vascular plants from which the other type originated in the course of evolution.
Brenner classified the protostele into following types
Haplostele: Protostele with smooth core of xylem surrounded by uniform layer of phloem is called haplostele. It is found in several fossil genera like Rynia and Horneophyton and living genera like Selaginella kraussiana.

Actinostele: a protostele in which xylem core is stellate or star shaped with radiating arms is called at actinostele. actor no still the phloem is not present in a continuous manner but in the form of separate groups which alternate with the arms of the star shaped xylem. Example Psilotum, Lycopodium serratum.

Sometimes the actinostele may show variations due to breaking of xylem tissue in two different forms. The common modifications of actinostele are
Plectostele: The xylem gets broken into a number of more or less parallel plates and such xylem plates alternate with phloem plates. Example Lycopodium clavatum and Lycopodium volubile.

Mixed protostele in this type of stele xylem appears in the form of irregular crops that are scared scattered in the ground mass of the phloem. E.g., L. cernuum.


Sometimes the actinostele may show variations due to breaking of xylem tissue in two different forms. The common modifications of actinostele are
Plectostele: The xylem gets broken into a number of more or less parallel plates and such xylem plates alternate with phloem plates. Example Lycopodium clavatum and Lycopodium volubile.

Mixed protostele in this type of stele xylem appears in the form of irregular crops that are scared scattered in the ground mass of the phloem. E.g., L. cernuum.

Siphonostele : It is a modified form of protostele in which the pith is present in the centre of the stele. A medulated protostele is called as siphonostele. During the development of siphonostele core of xylem is replaced by parenchymatous pith.
In siphonostele the central pith is surrounded by a cylinder of xylem and phloem. This type of stele is found in members of Pteropsida.
Origin of Siphonostele:
There is a general acceptance that the Siphonostele has evolved from protostele. Two theories have been proposed for evolution of Siphonostele from protostele,
I Extrastelar origin of pith: This theory was put forwarded by Jeffrey. According to him the pith originated as a result of the invasion of cortical parenchymatous cells into the stele. This invasion occurs through the leaf gaps and branch gaps.
II Intrastelar origin of pith:
Boodle, Gwynne-Vaughan, Bower supported this theory of intrastelar origin of pith.
According to this theory the innermost or centrally placed vascular tissue changes into parenchymatous cells which behave as pith. Presence of isolated trachieds in the pith in Botrychium viriginianum, Osmunda regalis support the intrastelar origin of pith.
Types of Siphonostele
Jeffrey classified the siphonostele into following two types on the basis of the position of phloem :
Ectophloic Siphonostele:
in this the earth is surrounded by xylem which in turn is surrounded by phloem, pericycle and endodermis. So in this case phloem is present only external to the xylem. E.g., Osmunda

Amphiphloic Siphonostele:
In this case the phloem is present on both the sides of xylem i.e., on external as well as internal sides. The pith is present in the centre. Xylem on inner side remains surrounded by inner phloem, inner pericycle, inner endodermis, and centrally placed pith. On the outer side of the xylem are present the outer phloem, outer pericycle and outer endodermis. It is found in Marsilea rhizome and Adiantum.

Modifications of siphonostele
Cladosiphonic siphonostele: A siphonostele with no leaf gap has been termed as clado siphonic siphonostele by Jeffrey. E.g., Selaginella.
Phyllosiphonic siphonostele: It is a siphonostele that remains perforated by smaller or larger leaf gaps. E.g., members of Fillicophyta.
The point at which the stem bundle diverges from the vascular cylinder toward the leaf is called as leaf gap..
If the stem or the rhizome bears leaves at distance the leaf gaps also appear at a considerable distance from each other. At such places the stele is interrupted and appears horseshoe shaped. The leaf gap closes at a higher level and the Stele again becomes complete and circular.
The siphonostele main assume different forms depending upon the occurrence, number and distribution of leaf gaps. These are as follows:
Solenostele: If the siphonostele is perforated by a single leaf gap only, corresponding to the origin of the leaf trace such a condition is known as solenostele. E
g., Adiantum pedatum.
Dictyostele: In advanced ferns like Dryopteris, Pteris the rhizome or stem is short and leaves overlap each other. This leads to the overlapping of leaf gaps in the stele so that the lower part of one leaf gap is parallel with the upper part of another leaf gap.
In Dictyostele the stele is broken into a network of seperate vascular strands, mainly because of crowded leaf gaps.
Each such separate vascular strand is known as meristele. Each meristele is of concentric type and consists of a central core of xylem surrounded by phloem, pericycle and endodermis.

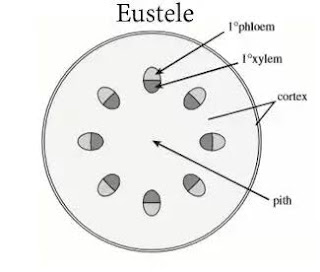
Eustele : if the stele is split into distinct collateral vascular bundle it is called eustele. It is a modification of the ectophloic siphonostele. E.g., Dicotyledons.
Atactostele: Differing from eustelic condition, the vascular strands are scattered. Such a stele has been named as atactostele by Esau. It occurs in monocotyledons.

Polycyclic stele: when the vascular tissue is present in the form of two or more concentric rings, such a stele is called a polycyclic stele. E.g., Pteridium aquilinium, Marattia.
Cladosiphonic siphonostele: A siphonostele with no leaf gap has been termed as clado siphonic siphonostele by Jeffrey. E.g., Selaginella.
Phyllosiphonic siphonostele: It is a siphonostele that remains perforated by smaller or larger leaf gaps. E.g., members of Fillicophyta.
The point at which the stem bundle diverges from the vascular cylinder toward the leaf is called as leaf gap..
If the stem or the rhizome bears leaves at distance the leaf gaps also appear at a considerable distance from each other. At such places the stele is interrupted and appears horseshoe shaped. The leaf gap closes at a higher level and the Stele again becomes complete and circular.
The siphonostele main assume different forms depending upon the occurrence, number and distribution of leaf gaps. These are as follows:
Solenostele: If the siphonostele is perforated by a single leaf gap only, corresponding to the origin of the leaf trace such a condition is known as solenostele. E
g., Adiantum pedatum.
Dictyostele: In advanced ferns like Dryopteris, Pteris the rhizome or stem is short and leaves overlap each other. This leads to the overlapping of leaf gaps in the stele so that the lower part of one leaf gap is parallel with the upper part of another leaf gap.
In Dictyostele the stele is broken into a network of seperate vascular strands, mainly because of crowded leaf gaps.
Each such separate vascular strand is known as meristele. Each meristele is of concentric type and consists of a central core of xylem surrounded by phloem, pericycle and endodermis.

Eustele : if the stele is split into distinct collateral vascular bundle it is called eustele. It is a modification of the ectophloic siphonostele. E.g., Dicotyledons.

Atactostele: Differing from eustelic condition, the vascular strands are scattered. Such a stele has been named as atactostele by Esau. It occurs in monocotyledons.
Polycyclic stele: when the vascular tissue is present in the form of two or more concentric rings, such a stele is called a polycyclic stele. E.g., Pteridium aquilinium, Marattia.




